Anatomy and Morphology
[This section contributed by Paula E. Cushing]
External Anatomy
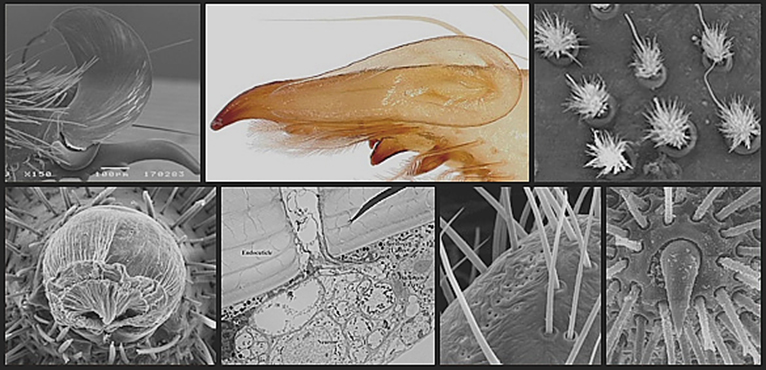
The “groundplan” morphology of the Solifugae has changed little since the Carboniferous (Petrunkevitch 1913, Selden & Dunlop 1998). Most solifuges are superficially similar in overall appearance; their morphology reflecting an errant, hunting lifestyle in the mostly arid environments where they are one of the dominant invertebrate predators. Nevertheless, there are interesting variations that have received little attention. The peculiar “mole solifuges” in the family Hexisopodidae (see Figure 1A) from southern Africa are highly modified for “swimming” through sand (Lamoral 1972, 1973). The compact Rhagodidae and Gylippidae are probably also fossorial (Figure 1B-C). Several southern African Solpugidae (Figure 1D) and Daesiidae are specialized for life on shifting sand dunes and display ecomorphological adaptations to this ultrapsammophilous habitat (Lawrence 1966, 1967, 1972). Some species of solifuges are specialist lithophiles, restricted to rocky scree-slopes (Wharton 1981).
FIGURE 1A-1F






Fig. 1A-1F: Representatives of six of the twelve families of Solifugae.
Fig. 1A: Hexisopodidae, photo by Colin Tilbury
Fig. 1B: Rhagodidae, photo by Chintan Sheth
Fig. 1C: Gylippidae, Gylippus ferganensis Birula, photo by Alexander Gromov
Fig. 1D: Solpugidae, Metasolpuga picta (Kraepelin), photo by EduVentures / Tharina Bird
Fig. 1E: Galeodidae, Paragaleodes pallidus (Birula), photo by Alexander Gromov
Fig. 1F: Eremobatidae, photo by Warren Savary.
Like most other arachnids, solifuges have a two-part body plan consisting of a prosoma, also called the peltidium, and an opisthosoma, also called the abdomen. The appendages (the chelicerae, legs, and pedipalps) are attached to the prosoma; the chelicerae are extraordinarily large (Figures 1E and F). The prosoma is, itself, divided into three sclerites: the propeltidium, the mesopeltidium, and the metapeltidium (Figure 2A).
FIGURE 2A
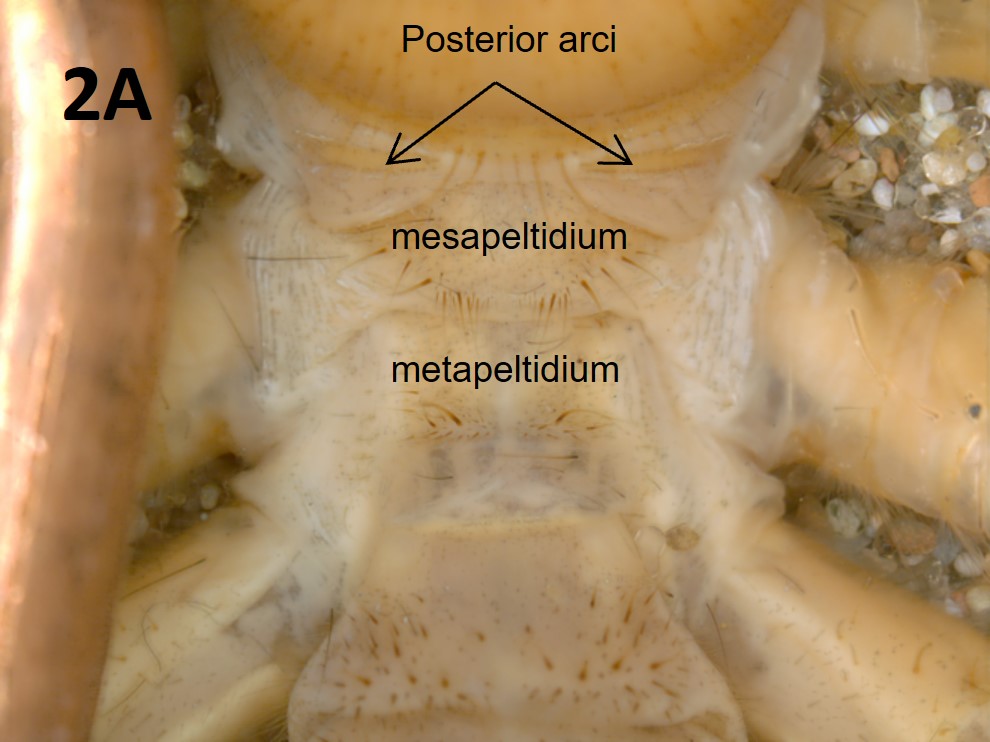
Fig. 2A: Tergites on the dorsum of Eremorhax joshui (Brookhart & Muma)
FIGURE 2B

Fig. 2B: Eremobates sp. (Eremobatidae) feeding to show the relative size of the chelicerae
Photo by Warren Savary
Two “ear-like” external lobes are attached to the anteriolateral edges of the propeltidum and give solifuges the appearance of having “Micky Mouse” ears. The anterior shape of the propeltidium (e.g., curved or straight) as well as the shape and apparent attachment to the propeltidium of the external lobes differs among the 12 families. Solifuges possess two simple eyes on a tubercle located medially along the anteriodorsal margin of the propeltidium (Figure 2B). The visual acuity of solifuges has not been tested; however, it is clear that they at least respond to light and dark (Graham et al. 2019).
One of the most apparent characteristics of solifuges is their large jaws, or chelicerae (Figures 1A-F and 2B). The chelicerae extend in front of the animal and consist of an immovable dorsal fixed finger and movable lower finger that articulates with the fixed finger (Figure 2C). In general, there is a single row of teeth on the ventral, movable finger and on the main part of the dorsal fixed finger. Adult males typically have reduced or modified dentition, especially on the upper fixed finger, relative to females and immatures. The pattern of dentition on male solifuge chelicerae is often diagnostic at the species level and the shape of the finger may also be modified. Male Eremobatidae typically lack dentition on the upper fixed cheliceral finger.
The powerful chelicerae are used to tear prey apart, dig burrows, threaten attackers (including solifuge biologists), and by males to transfer the sperm packet into the female’s gonopore (see Reproduction page of this website). A study by van der Meijden et al. (2012) measured the impressive bite force of two different species of solifuges. Solifuges in some families (e.g., Galeodidae) have stridulatory ridges located on the inner surfaces of the chelicerae. In some areas of the world, solifuges have been reported to make a hissing sound when disturbed, undoubtedly by rubbing these stridulatory ridges against one another (Pocock 1898, Cloudsley-Thompson 1961a, 1977; Cloudsley-Thompson & Constantinou 1984).
A beak or rostrum projects anteriorly between the two chelicerae. The rostrum includes the opening to the digestive tract (the mouth). The various plates of the rostrum are fused together, forming a lattice-like structure (Figure 2E; Klann & Alberti 2010). The entire rostrum acts to filter particulate material, limiting the animal’s diet primarily (but not entirely) to liquefied food (Hutton 1843; Turner 1916; Hingston 1925; Lawrence 1949; Muma 1966a). Bird et al. (2015) standardized the terminology for cheliceral characters and provided hypotheses of homology related to setal and tooth patterns.
FIGURE 2E

Fig. 2E: Rostrum located between chelicerae (filters particles).
Photo courtesy of Paula E. Cushing.
Males in several families have unique structures on the chelicerae termed flagella (Figures 2C and D), which are taxonomically diagnostic characters (Lamoral 1975; Bird et al. 2015). In some families (e.g., Melanoblossidae and Eremobatidae), the male flagellum consists of only modified setae and is not particularly conspicuous. In other families, the flagellum can be variously broad or narrow, membranous, or heavily sclerotized whip- or rod-like structures. In some taxa the flagella are fixed, while in others, they are paraxially moveable. Though used by at least some solifuges during spermatophore transfer (Wharton 1987; Peretti & Willemart 2007), the exact function of these often-bizarre structures remains unclear although various hypotheses have been proposed (Lamoral 1974).
FIGURE 2C

Fig. 2C: Flagellum complex of male Solpugiba lineata (C.L. Koch)
Photo by Alexander Gromov
FIGURE 2D

Fig. 2D: Flagellum complex of male Eusimonia divina Birula
Photo by Alexander Gromov
In addition to the chelicerae, solifuge appendages consist of often quite long, usually six-segmented pedipalps, a very delicate first pair of legs not used in locomotion, and three more pairs of locomotory legs. The pedipalps are typically held extended in front of the body when the solifuge is in motion and are frequently used to tap the ground while searching. Coxal glands (excretory organs) are found on the ventral surface of the pedipalps (Punzo 1998a). The pedipalps are covered in sensory setae that vary considerably in size and shape (Figure 2H; Roewer 1932). It is thought that the setae covering the pedipalps function, in general, as chemoreceptors, hygroreceptors, and thermoreceptors (Beccaloni 2009; Cushing & Casto 2012; Cushing et al. 2014); however, much work remains to be done to examine the structure and function of these various setae and other structures associated with solifuge pedipalps.
Suctorial organs are unique structures located at the tips of solifuge pedipalps (Figure 2F). These adhesive cuticular structures emerge, due to increased hemolymph pressure, from the tips of the palps only when their use is required; otherwise a simple set of muscles pull the suctorial organs back into the pedipalp and the tightly folded suctorial organ is covered by two cuticular plates (Cushing et al. 2005; Klann et al. 2008). The suctorial organs are thought to have evolved primarily for facilitating prey capture (Muma 1966a; Willemart et al. 2010) but are also used during mating, defense, and when the solifuge is climbing up a smooth surface (Cushing et al. 2005; Klann et al. 2008; Rowsell & Cushing 2020). In fact, Cushing et al. (2005) showed that the suctorial organs only are used when the solifuge is pulling itself up a smooth surface whereas the legs are used exclusive of the pedipalps when climbing up a rough surface.
FIGURE 2F
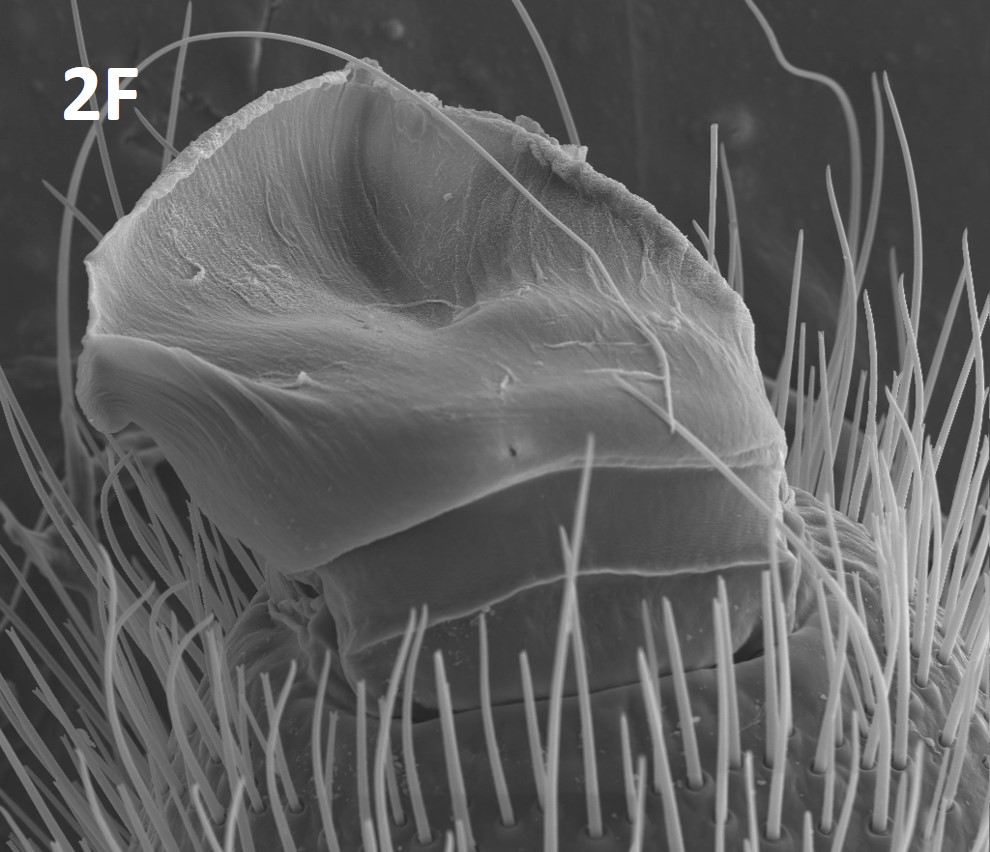
Fig. 2F: Suctorial organ of Eremorhax puebloensis
Photo by Brookhart, courtesy of Paula E. Cushing
The first pair of legs is typically shorter and more slender than the other appendages. In some species, the first legs are used in acquisition of food (Muma 1966b, 1967; Punzo 1998a; Hrušková-Martišová et al. 2008) although Wharton (1987) indicates that the first legs are not used in some Solpugidae. These slender legs are not used in locomotion; rather solifuges use the last three pairs for locomotion, and the 2nd pair is also the primary pair used when burrowing.
Unique fan-shaped structures called racquet organs, or malleoli, are found on the ventral surface of the fourth pair of legs (Figure 2G). There are typically 10 malleoli found on adults of both sexes. Juvenile solifuges have three to four pairs of malleoli (Muma 1966b; Klann 2009). Brownell & Farley (1974) performed histological and scanning electron microscopy (SEM) studies of these structures and determined that they serve as chemoreceptors, similar in function to the pectines found on scorpions.
FIGURE 2G

Fig. 2G: Malleoli of a solifuge
Photo by Paula E. Cushing
The abdomen of solifuges is 10 segmented. The external genital aperture (the gonopore) is located on the ventral side of the second opisthosomal segment. Compared with the epigynum of some spiders, the gonopore of female solifuges is relatively undifferentiated, although some differences in the genital plates surrounding the gonopore have been documented in numerous species descriptions (e.g., Brookhart 1965; Brookhart & Muma 1981, 1987; Brookhart & Cushing 2002, 2004, 2005; Catenazzi et al. 2009). During mating, males of Old World families release the sperm packet from their gonopores and then use their chelicerae to transfer the packet into the female gonopore (Amitai et al. 1962; Cloudsley-Thompson 1961b, 1977; Junqua 1966; Heymons 1902; Wharton 1987); in the New World Eremobatidae, males deposit the sperm packet directly onto the female gonopore from their own after which they use their chelicerae to break open the sperm packet (Muma 1966c). See Reproduction for more information.
Internal Anatomy
Solifuges have three pair of spiracles opening into the extensive tracheal systems: one pair is located on the prosoma, posterior to the coxal joint of the 2nd pair of legs; and two pair on the 2nd, 3rd, or sometimes 4th opisthosomal segments respectively (Babák 1921; Nogge 1976; Anja Klann pers. obs.). The tracheael system in solifuges is extensive, appears to have evolved convergently from the tracheae of insects, but provides solifuges with an efficient gas exchange system that allows them to have a higher metabolic and respiratory rate more similar to insects than to other arachnids (Lighton & Fielden 1996; Franz-Guess et al. 2016). The abdominal spiracles are often associated with stiff spine-like setae called ctenidia (pers. obs). Perhaps as a result of their extensive tracheal system, solifuges are able to run faster and for more prolonged periods than most other arachnids (Wharton 1987; Punzo 1998c; Ken Prestwich pers. comm.). Most solifuges are highly active cursorial predators, and an efficient system for oxygen intake, such as that provided by the trachea, is essential for this mode of life. The anus is located terminally on the abdomen except in the family Rhagodidae, in which it is ventrally located.
Modern new methods and techniques are beginning to reveal remarkable new structures along with their purported function (Barth & Stagl 1976; Alberti 1979; Haupt 1982; Bauchhenss 1983; Cloudsley-Thompson & Constantinou 1984; Klann et al. 2005; Klann & Alberti 2010), yet much work remains to be done to elucidate the structural significance of solifuge anatomy.